Нейрон - электрически возбудимая клетка, которая обрабатывает, хранит и передает информацию с помощью электрических и химических сигналов. Клетка содержит ядро, тело клетки и отростки (дендриты и аксоны). В головном мозге человека насчитывается в среднем около 65 миллиардов нейронов. Нейроны соединяются между собой, формируя таким образом человеческие функции мозга, память, отделы и сознание.
Видите это изображение выше? С помощью этого странного изображения нейробиологи Массачусетского технологического института смогли активировать отдельные нейроны мозга. Используя лучшую из доступных модель зрительной нейронной сети мозга, ученые разработали новый способ точного управления отдельными нейронами и их популяциями в середине этой сети. В ходе испытания на животных команда показала, что информация, полученная из вычислительной модели, позволила им создавать изображения, которые сильно активировали определенные нейроны мозга.
Свежий вкусен сам по себе, из винограда делают замечательные , а если высушить виноград — получится сладкий изюм. Но какие ещё плюсы несёт в себе эта ягода с богатейшей историей? Исследователи из Школы медицины на горе Синай пришли к выводу, что на основе винограда можно создать эффективное и безопасное средство против депрессии, которое будет обладать минимальными побочными эффектами для здоровья человека.
Была представлена модель нервной системы, опишу теорию и принципы, которые легли в её основу.
Теория основана на анализе имеющейся информации о биологическом нейроне и нервной системе из современной нейробиологии и физиологии мозга.
Сначала приведу краткую информацию об объекте моделирования, вся информация изложена далее, учтена и использована в модели.
НЕЙРОН
Нейрон является основным функциональным элементом нервной системы, он состоит из тела нервной клетки и её отростков. Существуют два вида отростков: аксоны и дендриты. Аксон – длинный покрытый миелиновой оболочкой отросток, предназначенный для передачи нервного импульса на далекие расстояния. Дендрит – короткий, ветвящийся отросток, благодаря которым происходит взаимосвязь с множеством соседних клеток.
ТРИ ТИПА НЕЙРОНОВ
Нейроны могут сильно отличаться по форме, размерам и конфигурации, не смотря на это, отмечается принципиальное сходство нервной ткани в различных участках нервной системе, отсутствуют и серьезные эволюционные различия. Нервная клетка моллюска Аплизии может выделять такие же нейромедиаторы и белки, что и клетка человека.В зависимости от конфигурации выделяют три типа нейронов:
А) рецепторные, центростремительные, или афферентные нейроны, данные нейроны имеют центростремительный аксон, на конце которого имеются рецепторы, рецепторные или афферентные окончания. Эти нейроны можно определить, как элементы, передающие внешние сигналы в систему.
Б) интернейроны (вставочные, контактные, или промежуточные) нейроны, не имеющие длинных отростков, но имеющие только дендриты. Таких нейронов в человеческом мозгу больше чем остальных. Данный вид нейронов является основным элементом рефлекторной дуги.
В) моторные, центробежные, или эфферентные, они имеют центростремительный аксон, который имеет эфферентные окончания передающий возбуждение мышечным или железистым клеткам. Эфферентные нейроны служат для передачи сигналов из нервной среды во внешнюю среду.
Обычно в статьях по искусственным нейронным сетям оговаривается наличие только моторных нейронов (с центробежным аксоном), которые связаны в слои иерархической структуры. Подобное описание применимо к биологической нервной системе, но является своего рода частным случаем, речь идет о структурах, базовых условных рефлексов. Чем выше в эволюционном значении нервная система, тем меньше в ней превалируют структуры типа «слои» или строгая иерархия.
ПЕРЕДАЧА НЕРВНОГО ВОЗБУЖДЕНИЯ
Передача возбуждения происходит от нейрона к нейрону, через специальные утолщения на концах дендритов, называемых синапсами. По типу передачи синапсы разделяют на два вида: химические и электрические. Электрические синапсы передают нервный импульс непосредственно через место контакта. Таких синапсов в нервных системах очень мало, в моделях не будут учитываться. Химические синапсы передают нервный импульс посредством специального вещества медиатора (нейромедиатора, нейротрансмиттера), данный вид синапса широко распространен и подразумевает вариативность в работе.Важно отметить, что в биологическом нейроне постоянно происходят изменения, отращиваются новые дендриты и синапсы, возможны миграции нейронов. В местах контактов с другими нейронами образуются новообразования, для передающего нейрона - это синапс, для принимающего - это постсинаптическая мембрана, снабжаемая специальными рецепторами, реагирующими на медиатор, то есть можно говорить, что мембрана нейрона - это приемник, а синапсы на дендритах - это передатчики сигнала.
СИНАПС

При активации синапса он выбрасывает порции медиатора, эти порции могут варьироваться, чем больше выделится медиатора, тем вероятнее, что принимаемая сигнал нервная клетка будет активирована. Медиатор, преодолевая синоптическую щель, попадает на постсинаптическую мембрану, на которой расположены рецепторы, реагирующие на медиатор. Далее медиатор может быть разрушен специальным разрушающим ферментом, либо поглощен обратно синапсом, это происходит для сокращения времени действия медиатора на рецепторы.
Так же помимо побудительного воздействия существуют синапсы, оказывающие тормозящее воздействие на нейрон. Обычно такие синапсы принадлежат определенным нейронам, которые обозначаются, как тормозящие нейроны.
Синапсов связывающих нейрон с одной и той же целевой клеткой, может быть множество. Для упрощения примем, всю совокупность, оказываемого воздействия одним нейроном, на другой целевой нейрон за синапс с определённой силой воздействия. Главной характеристикой синапса будет, является его сила.
СОСТОЯНИЕ ВОЗБУЖДЕНИЯ НЕЙРОНА
В состоянии покоя мембрана нейрона поляризована. Это означает, что по обе стороны мембраны располагаются частицы, несущие противоположные заряды. В состоянии покоя наружная поверхность мембраны заряжена положительно, внутренняя – отрицательно. Основными переносчиками зарядов в организме являются ионы натрия (Na+), калия (K+) и хлора (Cl-).Разница между зарядами поверхности мембраны и внутри тела клетки составляет мембранный потенциал. Медиатор вызывает нарушения поляризации – деполяризацию. Положительные ионы снаружи мембраны устремляются через открытые каналы в тело клетки, меняя соотношение зарядов между поверхностью мембраны и телом клетки.

Изменение мембранного потенциала при возбуждении нейрона
Характер изменений мембранного потенциала при активации нервной ткани неизменен. Независимо от того кокой силы воздействия оказывается на нейрон, если сила превышает некоторое пороговое значение, ответ будет одинаков.
Забегая вперед, хочу отметить, что в работе нервной системы имеет значение даже следовые потенциалы (см. график выше). Они не появляются, вследствие каких-то гармонических колебаний уравновешивающих заряды, являются строгим проявлением определённой фазы состояния нервной ткани при возбуждении.
ТЕОРИЯ ЭЛЕКТРОМАГНИТНОГО ВЗАИМОДЕЙСТВИЯ
Итак, далее приведу теоретические предположения, которые позволят нам создавать математические модели. Главная идея заключается во взаимодействии между зарядами формирующихся внутри тела клетки, во время её активности, и зарядами с поверхностей мембран других активных клеток. Данные заряды являются разноименными, в связи этим можно предположить, как будут располагаться заряды в теле клетки под воздействием зарядов других активных клеток.
Можно сказать, что нейрон чувствует активность других нейронов на расстоянии, стремится направить распространения возбуждения в направлении других активных участков.
В момент активности нейрона можно рассчитать определённую точку в пространстве, которая определялась бы, как сумма масс зарядов, расположенных на поверхностях других нейронов. Указанную точку назовем точкой паттерна, её месторождение зависит от комбинации фаз активности всех нейронов нервной системы. Паттерном в физиологии нервной системы называется уникальная комбинация активных клеток, то есть можно говорить о влиянии возбуждённых участков мозга на работу отдельного нейрона.
Нужно представлять работу нейрона не просто как вычислителя, а своего рода ретранслятор возбуждения, который выбирает направления распространения возбуждения, таким образом, формируются сложные электрические схемы. Первоначально предполагалось, что нейрон просто избирательно отключает/включает для передачи свои синапсы, в зависимости от предпочитаемого направления возбуждения. Но более детальное изучение природы нейрона, привело к выводам, что нейрон может изменять степень воздействия на целевую клетку через силу своих синапсов, что делает нейрон более гибким и вариативным вычислительным элементом нервной системы.

Какое же направление для передачи возбуждения является предпочтительным? В различных экспериментах связанных с образованием безусловных рефлексов, можно определить, что в нервной системе образуются пути или рефлекторные дуги, которые связывают активируемые участки мозга при формировании безусловных рефлексов, создаются ассоциативные связи. Значит, нейрон должен передавать возбуждения к другим активным участкам мозга, запоминать направление и использовать его в дальнейшем.
Представим вектор начало, которого находится в центре активной клети, а конец направлен в точку паттерна определённую для данного нейрона. Обозначим, как вектор предпочитаемого направления распространения возбуждения (T, trend). В биологическом нейроне вектор Т может проявляться в структуре самой нейроплазмы, возможно, это каналы для движения ионов в теле клетки, или другие изменения в структуре нейрона.
Нейрон обладает свойством памяти, он может запоминать вектор Т, направление этого вектора, может меняться и перезаписываться в зависимости от внешних факторов. Степень с которой вектор Т может подвергается изменениям, называется нейропластичность.
Этот вектор в свою очередь оказывает влияние на работу синапсов нейрона. Для каждого синапса определим вектор S начало, которого находится в центре клетки, а конец направлен в центр целевого нейрона, с которым связан синапс. Теперь степень влияния для каждого синапса можно определить следующим образом: чем меньше угол между вектором T и S, тем больше синапс будет, усиливается; чем меньше угол, тем сильнее синапс будет ослабевать и возможно может прекратить передачу возбуждения. Каждый синапс имеет независимое свойство памяти, он помнит значение своей силы. Указанные значения изменяются при каждой активизации нейрона, под влиянием вектора Т, они либо увеличиваются, либо уменьшаются на определённое значение.
МАТЕМАТИЧЕСКАЯ МОДЕЛЬ

Входные сигналы (x1, x2,…xn) нейрона представляют собой вещественные числа, которые характеризуют силу синапсов нейронов, оказывающих воздействие на нейрон.
Положительное значение входа означает побудительное воздействие, оказываемое на нейрон, а отрицательное значение – тормозящее воздействие.
Для биологического нейрона не имеет значение, откуда поступил возбуждающий его сигнал, результат его активности будет идентичен. Нейрон будет активизирован, когда сумма воздействий на него будет превышать определённое пороговое значение. Поэтому, все сигналы проходят через сумматор (а), а поскольку нейроны и нервная система работают в реальном времени, следовательно, воздействие входов должно оцениваться в короткий промежуток времени, то есть воздействие синапса имеет временный характер.
Результат сумматора проходит пороговую функцию (б), если сумма превосходит пороговое значение, то это приводит к активности нейрона.
При активации нейрон сигнализирует о своей активности системе, передовая информацию о своём положении в пространстве нервной системы и заряде, изменяемом во времени (в).
Через определённое время, после активации нейрон передает возбуждение по всем имеющимся синапсам, предварительно производя пересчет их силы. Весь период активации нейрон перестает реагировать на внешние раздражители, то есть все воздействия синапсов других нейронов игнорируются. В период активации входит так же период восстановления нейрона.
Происходит корректировка вектора Т (г) с учётом значения точки паттерна Pp и уровнем нейропластичности. Далее происходит переоценка значений всех сил синапсов в нейроне(д).
Обратите внимание, что блоки (г) и (д) выполняются параллельно с блоком (в).
ЭФФЕКТ ВОЛНЫ
Если внимательно проанализировать предложенную модель, то можно увидеть, что источник возбуждения должен оказывать большее влияние на нейрон, чем другой удалённый, активный участок мозга. Следовательно возникает вопрос: почему же все равно происходит передача в направлении другого активного участка?Данную проблему я смог определить, только создав компьютерную модель. Решение подсказал график изменения мембранного потенциала при активности нейрона.

Усиленная реполяризация нейрона, как говорилось ранее, имеет важное значение для нервной системы, благодаря ей создается эффект волны, стремление нервного возбуждения распространятся от источника возбуждения.
При работе с моделью я наблюдал два эффекта, ели пренебречь следовым потенциалом или сделать его недостаточно большим, то возбуждение не распространяется от источников, а в большей степени стремится к локализации. Если сделать следовой потенциал сильно большим, то возбуждение стремится «разбежаться» в разные стороны, не только от своего источника, но и от других.
КОГНИТИВНАЯ КАРТА
Используя теорию электромагнитного взаимодействия, можно объяснить многие явления и сложные процессы, протекающие в нервной системе. К примеру, одним из последних открытий, которое широко обсуждается в науках о мозге, является открытие когнитивных карт в гиппокампе.Гиппокамп – это отдел мозга, которому отвечает за кратковременную память. Эксперименты на крысах выявили, что определённому месту в лабиринте соответствует своя локализованная группа клеток в гиппокампе, причем, не имеет значение, как животное попадает в это место, все равно будет активирован соответствующий этому месту участок нервной ткани. Естественно, животное должно помнить данный лабиринт, не стоит рассчитывать на топологическое соответствие пространства лабиринта и когнитивной карты.

Каждое место в лабиринте представляется в мозге, как совокупность раздражителей различного характера: запахи, цвет стен, возможные примечательные объекты, характерные звуки и т. д. Указанные раздражители отражаются на коре, различных представительствах органов чувств, в виде всплесков активности в определённых комбинациях. Мозг одновременно обрабатывает информацию в нескольких отделах, зачастую информационные каналы разделяются, одна и та же информация поступает в различные участки мозга.

Активация нейронов места в зависимости от положения в лабиринте (активность разных нейронов показана разным цветом).
Гиппокамп расположен в центре мозга, вся кара и её области удалены от него, на одинаковые расстояния. Если определить для каждой уникальной комбинации раздражителей точку масс зарядов поверхностей нейронов, то можно увидеть, что указанные точки будут различны, и будут находиться примерно в центре мозга. К этим точкам будет стремиться и распространятся возбуждение в гиппокампе, формируя устойчивые участки возбуждения. Более того, поочередная смена комбинаций раздражителей, будет приводить к смещению точки паттерна. Участки когнитивной карты будут ассоциативно связываться друг с другом последовательно, что приведет к тому, что животное, помещенное в начало знакомого ей лабиринта, может вспомнить весь последующий путь.
Заключение
У многих возникнет вопрос, где в данной работе предпосылки к элементу разумности или проявления высшей интеллектуальной деятельности?Важно отметить, что феномен человеческого поведения, есть следствие функционирования биологической структуры. Следовательно, чтобы имитировать разумное поведение, необходимо хорошо понимать принципы и особенности функционирования биологических структур. К сожалению, в науке биологии пока не представлен четкий алгоритм: как работает нейрон, как понимает, куда необходимо отращивать свои дендриты, как настроить свои синапсы, что бы в нервной системе смог сформироваться простой условный рефлекс, на подобие тех, которые демонстрировал и описывал в своих работах академик И.П. Павлов.
С другой стороны в науке об искусственном интеллекте, в восходящем (биологическом) подходе, сложилось парадоксальная ситуация, а именно: когда используемые в исследованиях модели основаны на устаревших представлениях о биологическом нейроне, консерватизм, в основе которого берётся персептрон без переосмысления его основных принципов, без обращения к биологическому первоисточнику, придумывается все более хитроумные алгоритмы и структуры, не имеющих биологических корней.
Конечно, никто не уменьшает достоинств классических нейронных сетей, которые дали множество полезных программных продуктов, но игра с ними не является путем к созданию интеллектуально действующей системы.
Более того, не редки заявления, о том, что нейрон подобен мощной вычислительной машине, приписывают свойство квантовых компьютеров. Из-за этой сверхсложности, нервной системе приписывается невозможность её повторения, ведь это соизмеримо с желанием смоделировать человеческую душу. Однако, в реальности природа идет по пути простоты и элегантности своих решений, перемещение зарядов на мембране клетки может служить, как для передачи нервного возбуждения, так и для трансляции информации о том, где происходит данная передача.
Несмотря на то, что указанная работа демонстрирует, как образуются элементарные условные рефлексы в нервной системе, она приближает к пониманию того, что такое интеллект и разумная деятельность.
Существуют еще множество аспектов работы нервной системы: механизмы торможения, принципы построения эмоций, организация безусловных рефлексов и обучение, без которых невозможно построить качественную модель нервной системы. Есть понимание, на интуитивном уровне, как работает нервная система, принципы которой возможно воплотить в моделях.
Создание первой модели помогли отработать и откорректировать представление об электромагнитном взаимодействии нейронов. Понять, как происходит формирование рефлекторных дуг, как каждый отдельный нейрон понимает, каким образом ему настроить свои синапсы для получения ассоциативных связей.
На данный момент я начал разрабатывать новую версию программы, которая позволит смоделировать многие другие аспекты работы нейрона и нервной системы.
Прошу принять активное участие в обсуждении выдвинутых здесь гипотез и предположений, так как я могу относиться к своим идеям предвзято. Ваше мнение очень важно для меня.
Теги: Добавить метки
Отдельные нервные клетки, или нейроны , выполняют свои функции не как изолированные единицы, подобно клеткам печени или почек. Работа 50 миллиардов (или около того) нейронов нашего мозга состоит в том, что они получают сигналы от каких-то других нервных клеток и передают их третьим.
Передающие и принимающие клетки объединены в нервные цепи или сети (см. рис. 26). Отдельный нейрон с дивергентной структурой (от лат. diverge - отклоняюсь) может посылать сигналы тысяче и даже большему числу других нейронов. Но чаще один такой нейрон соединяется всего лишь с несколькими определенными нейронами. Точно так же какой-либо нейрон может получать входную информацию от других нейронов с помощью одной, нескольких или многих входных связей, если на нем сходятся конвергентные пути (от лат. converge - приближаюсь, схожусь). Конечно, все зависит от того, какую именно клетку мы рассматриваем и в какую сеть она оказалась включенной в процессе развития. Вероятно, в каждый момент времени активна лишь небольшая часть путей, оканчивающихся на данном нейроне.
Действительные места соединения - специфические точки на поверхности нервных клеток, где происходит их контакт, - называются синапсами (synapsis; греч. «соприкосновение», «соединение») (см. рис. 26 и 27), а сам процесс передачи информации в этих местах - синаптической передачей . При взаимодействии нейронов с помощью синаптической передачи посылающая сигнал (пресинаптическая) клетка выделяет определенное вещество на рецепторную поверхность воспринимающего (постсинаптического) нейрона. Это вещество, называемое нейромедиатором , служит молекулярным посредником для передачи информации от передающей клетки к воспринимающей. Нейромедиатор замыкает цепь, осуществляя химическую передачу информации через синаптическую щель - структурный разрыв между передающей и воспринимающей клетками в месте синапса.
Особенности нервных клеток
Нейроны обладают рядом признаков, общих для всех клеток тела. Независимо от своего местонахождения и функций любой нейрон, как и всякая другая клетка, имеет плазматическую мембрану , определяющую границы индивидуальной клетки. Когда нейрон взаимодействует с другими нейронами или улавливает изменения в локальной среде, он делает это с помощью плазматической мембраны и заключенных в ней молекулярных механизмов.
Все, что находится внутри плазматической мембраны (кроме ядра), называется цитоплазмой . Здесь содержатся цитоплазматические органеллы , необходимые для существования нейрона и выполнения им своей работы (см. рис. 27 и 28). Митохондрии обеспечивают клетку энергией, используя сахар и кислород для синтеза специальных высокоэнергетических молекул, расходуемых клеткой по мере надобности. Микротрубочки - тонкие опорные структуры - помогают нейрону сохранять определенную форму. Сеть внутренних мембранных канальцев, с помощью которых клетка распределяет продукты, необходимые для ее функционирования, называется эндоплазматическим ретикуломом .
Существует два вида эндоплазматического ретикулума. Мембраны «шероховатого», или гранулярного, ретикулума усеяны рибосомами , необходимыми клетке для синтеза секретируемых ею белковых веществ. Обилие элементов шероховатого ретикулума в цитоплазме нейронов характеризует их как клетки с весьма интенсивной секреторной деятельностью. Белки, предназначенные только для внутриклеточного использования, синтезируются на многочисленных рибосомах, не прикрепленных к мембранам ретикулума, а находящихся в цитоплазме в свободном состоянии. Другой вид эндоплазматического ретикулума называют «гладким». Органеллы, построенные из мембран гладкого ретикулума, упаковывают продукты, предназначенные для секреции, в «мешочки» из таких мембран для последующего переноса их к поверхности клетки, где они выводятся наружу. Гладкий эндоплазматический ретикулум называют также аппаратом Гольджи , по имени итальянца Эмилио Гольджи, который впервые разработал метод окраски этой внутренней структуры, сделавший возможным ее микроскопическое изучение.
Камилло Гольджи (1844-1926). Фотография сделана в начале 1880-х годов, когда Гольджи был профессором университета в Павии. В 1906 году он разделил с Кахалом Нобелевскую премию по физиологии и медицине.

Сантьяго Рамон-и-Кахал (1852-1934). Поэт, художник и гистолог, обладавший поразительным творческим потенциалом, он преподавал в основном в Мадридском университете. Этот автопортрет он создал в 1920-х годах.
В центре цитоплазмы находится клеточное ядро . Здесь у нейронов, как и у всех клеток с ядрами, содержится генетическая информация, закодированная в химической структуре генов. В соответствии с этой информацией полностью сформированная клетка синтезирует специфические вещества, которые определяют форму, химизм и функции этой клетки. В отличие от большинства других клеток тела зрелые нейроны не могут делиться, и генетически обусловленные продукты любого нейрона должны обеспечивать сохранение и изменение его функций на протяжении всей его жизни.
Нейроны сильно различаются по своей форме, по связям, которые они образуют, и способам функционирования. Наиболее очевидное отличие нейронов от других клеток заключается в разнообразии их размеров и формы. Большинство клеток тела имеет шарообразную, кубическую или пластинчатую форму. Для нейронов же характерны неправильные очертания: у них имеются отростки, часто многочисленные и разветвленные. Эти отростки - живые «провода», с помощью которых образуются нейронные цепи. Нервная клетка имеет один главный отросток, называемый аксоном (греч. ax?n - ось), по которому она передает информацию следующей клетке в нейронной цепи. Если нейрон образует выходные связи с большим числом других клеток, его аксон многократно ветвится, чтобы сигналы могли дойти до каждой из них.

Рис. 28. Внутреннее строение типичного нейрона. Микротрубочки обеспечивают структурную жесткость, а также транспортировку материалов, синтезируемых в теле клетки и предназначенных для использования в окончании аксона (внизу). В этом окончании находятся синаптические пузырьки, содержащие медиатор, а также пузырьки, выполняющие иные функции. На поверхности постсинаптического дендрита показаны предполагаемые места рецепторов для медиатора (см. также рис. 29).
Другие отростки нейрона называются дендритами . Этот термин, происходящий от греческого слова dendron - «дерево», означает, что они имеют древовидную форму. На дендритах и на поверхности центральной части нейрона, окружающей ядро (и называемой перикарионом , или телом клетки), находятся входные синапсы, образуемые аксонами других нейронов. Благодаря этому каждый нейрон оказывается звеном той или иной нейронной сети.
В разных участках цитоплазмы нейрона содержатся различные наборы специальных молекулярных продуктов и органелл. Шероховатый эндоплазматический ретикулум и свободные рибосомы обнаружены только в цитоплазме тела клетки и в дендритах. В аксонах эти органеллы отсутствуют, и поэтому синтез белка здесь невозможен. Окончания аксонов содержат органеллы, называемые синаптическими пузырьками , в которых находятся молекулы медиатора, выделяемого нейроном. Полагают, что каждый синаптический пузырек несет в себе тысячи молекул вещества, которое используется нейроном для передачи сигналов другим нейронам (см. рис. 29).

Рис. 29. Схема выброса медиатора и процессов, происходящих в гипотетическом центральном синапсе.
Дендриты и аксоны сохраняют свою форму благодаря микротрубочкам, которые, по-видимому, играют также роль в передвижении синтезированных продуктов из центральной цитоплазмы к очень далеким от нее концам ветвящихся аксонов и дендритов. При методе окраски, разработанном Гольджи, используется металлическое серебро, которое связывается с микротрубочками и позволяет выявить форму изучаемой нервной клетки. В начале XX века испанский микроанатом Сантьяго Рамон-и-Кахал почти интуитивно применил этот метод для установления клеточной природы организации мозга и для классификации нейронов в соответствии с их уникальными и общими структурными особенностями.
Различные названия нейронов
Нейроны в зависимости от контекста могут называться по-разному. Иногда это может сбить с толку, но на самом деле это очень похоже на то, как мы называем себя или наших знакомых. Смотря по обстоятельствам, мы говорим об одной и той же девушке как о студентке, дочери, сестре, рыжеволосой красавице, пловчихе, любимой или члене семейства Смитов. Нейроны тоже получают столько ярлыков, сколько различных ролей они выполняют. Разные ученые использовали, вероятно, все достойные внимания свойства нейронов в качестве основы для их классификации.
Каждая уникальная структурная особенность того или иного нейрона отражает степень его специализации для выполнения определенных задач. Можно называть нейроны в соответствии с этими задачами, или функциями. Это один способ. Например, нервные клетки, объединенные в цепи, которые помогают нам воспринимать внешний мир или контролировать события, происходящие внутри нашего тела, именуются сенсорными (чувствительными) нейронами. Нейроны, объединенные в сети, вызывающие мышечные сокращения и, следовательно, движение тела, называются моторными или двигательными.
Положение нейрона в сети - другой важный критерий наименования. Нейроны, ближе всего расположенные к месту действия (будь то ощущаемый стимул или активируемая мышца), - это первичные сенсорные или моторные нейроны, или нейроны первого порядка. Далее следуют вторичные нейроны (нейроны второго порядка), затем третичные (третьего порядка) и т.д.
Регуляция нейронной активности
Способность нервной системы и мышц генерировать электрические потенциалы известна давно - со времен работ Гальвани в конце XVIII столетия. Однако наши знания о том, как возникает это биологическое электричество при функционировании нервной системы, основаны на исследованиях всего лишь 25-летней давности.
Все живые клетки обладают свойством «электрической полярности». Это означает, что по отношению к какой-то отдаленной и явно нейтральной точке (электрики называют ее «землей») внутренняя часть клетки испытывает относительный недостаток положительно заряженных частиц и поэтому, как мы говорим, отрицательно заряжена относительно наружной стороны клетки. Что же это за частицы, находящиеся внутри и вне клеток нашего тела?
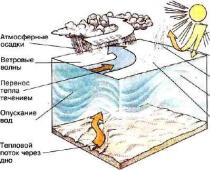
Жидкости нашего тела - плазма, в которой плавают клетки крови, внеклеточная жидкость, заполняющая пространство между клетками различных органов, спинномозговая жидкость, находящаяся в желудочках мозга, - все это особые разновидности соленой воды. (Некоторые ученые, мыслящие историческими категориями, усматривают в этом следы того периода эволюции, когда все живые создания существовали в первичном океане.) Встречающиеся в природе соли обычно составлены из нескольких химических элементов - натрия, калия, кальция и магния, несущих положительные заряды в жидкостях тела, и хлорида, фосфата и остатков некоторых более сложных кислот, образуемых клетками и несущих отрицательный заряд. Заряженные молекулы или атомы именуются ионами .
Во внеклеточных пространствах положительные и отрицательные ионы распределены свободно и в равных количествах, так что они нейтрализуют друг друга. Внутри клеток, однако, относительный дефицит положительно заряженных ионов обусловливает общий отрицательный заряд. Этот отрицательный заряд возникает потому, что плазматическая мембрана проницаема не для всех солей в равной мере. Некоторые ионы, например К + , обычно проникают сквозь мембрану легче, чем другие, такие, как ионы натрия (Na +) или кальция (Ca 2+). Внеклеточные жидкости содержат довольно много натрия и мало калия. Внутри клеток жидкости относительно бедны натрием и богаты калием, но общее содержание положительных ионов внутри клетки не вполне уравновешивает отрицательные заряды хлорида, фосфата и органических кислот цитоплазмы. Калий проходит через клеточную мембрану лучше других ионов и, по-видимому, весьма склонен выходить наружу, так как концентрация его внутри клеток намного выше, чем в окружающей их среде. Таким образом, распределение ионов и избирательность их перехода через полупроницаемую мембрану приводят к созданию отрицательного заряда внутри клеток.
В то время как описанные факторы ведут к установлению трансмембранной ионной полярности, другие биологические процессы способствуют ее поддержанию. Один из таких факторов - очень эффективные ионные насосы, которые существуют в плазматической мембране и получают энергию от митохондрий. Такие насосы «откачивают» ионы натрия, поступающие в клетку с молекулами воды или сахара.
«Электрически возбудимые» клетки, подобные нейронам, обладают способностью регулировать свой внутренний отрицательный потенциал. При воздействии некоторых веществ в «возбуждающих» синапсах свойства плазматической мембраны постсинаптического нейрона изменяются. Внутренность клетки начинает терять свой отрицательный заряд, и натрий не встречает больше препятствий для перехода внутрь через мембрану. Действительно, после проникновения в клетку некоторого количества натрия переход натрия и других положительных ионов (кальция и калия) внутрь клетки, т.е. деполяризация, во время краткого периода возбуждения протекает столь успешно, что внутренность нейрона менее чем на 1/1000 секунды становится заряженной положительно. Этот переход от обычного отрицательного состояния содержимого клетки к кратковременному положительному называют потенциалом действия или нервным импульсом . Положительное состояние длится так недолго потому, что реакция возбуждения (повышенное поступление в клетку натрия) носит саморегулируемый характер. Присутствие повышенных количеств натрия и кальция в свою очередь ускоряет эвакуацию калия, по мере того как ослабевает действие возбуждающего импульса. Нейрон быстро восстанавливает электрохимическое равновесие и возвращается к состоянию с отрицательным потенциалом внутри до следующего сигнала.

Рис. 30. Когда нейрон активируется приходящим к нему возбуждающим импульсом, волна деполяризации временно меняет знак мембранного потенциала. По мере распространения волны деполяризации вдоль аксона последовательные участки аксона тоже претерпевают эту временную реверсию. Потенциал действия можно описать как поток положительно заряженных ионов натрия (Na +), переходящих через мембрану внутрь нейрона.
Деполяризация, связанная с потенциалом действия, распространяется вдоль аксона как волна активности (рис. 30). Движение ионов, возникающее около деполяризованного участка, способствует деполяризации следующего участка, и в результате каждая волна возбуждения быстро достигает всех синаптических окончаний аксона. Главное преимущество электрического проведения импульса по аксону состоит в том, что возбуждение быстро распространяется на большие расстояния без какого-либо ослабления сигнала.
Кстати, нейроны с короткими аксонами, по-видимому, не всегда генерируют нервные импульсы. Это обстоятельство, если оно будет твердо установлено, может иметь далеко идущие последствия. Если клетки с короткими аксонами способны изменять уровень активности, не генерируя потенциалы действия, то исследователи, пытающиеся по электрическим разрядам оценить роль отдельных нейронов в определенных видах поведения, легко могут упустить из виду многие из важных функций «молчащих» клеток.
Синаптические медиаторы
С некоторыми оговорками синапсы можно сравнить с перекрестками на проводящих путях мозга. В синапсах сигналы передаются только в одном направлении - с концевой веточки посылающего их пресинаптического нейрона на ближайший участок постсинаптического нейрона. Однако быстрая электрическая передача, так хорошо действующая в аксоне, в синапсе не работает. Не вдаваясь в биологические причины этого, мы можем просто констатировать, что химическая связь в синапсах обеспечивает более тонкую регуляцию свойств мембраны постсинаптической клетки.
При общении друг с другом люди передают основное содержание своей речи словами. Чтобы сделать более тонкие акценты или подчеркнуть дополнительный смысл слов, они пользуются тембром голоса, мимикой, жестами. При коммуникации нервных клеток основные единицы информации передаются специфическими химическими посредниками - синаптическими медиаторами (определенный нейрон использует один и тот же медиатор во всех своих синапсах). Если продолжить нашу аналогию с вербальным и невербальным способом общения, то можно сказать, что одни химические посредники передают «факты», а другие - дополнительные смысловые оттенки или акценты.

Рис. 31. Противоположное действие возбуждающего (слева) и тормозного (справа) медиаторов можно объяснить тем, что они влияют на разные ионные каналы.
Вообще говоря, существуют два вида синапсов - возбуждающие и тормозные (рис. 31). В первом случае одна клетка приказывает другой переходить к активности, а во втором, наоборот, затрудняет активацию клетки, которой передается сигнал. Под действием постоянных тормозящих команд некоторые нервные клетки хранят молчание до тех пор, пока возбуждающие сигналы не заставят их активироваться. Например, нервные клетки спинного мозга, которые приказывают вашим мышцам действовать, когда вы ходите или танцуете, обычно «молчат», пока не получат возбуждающих импульсов от клеток моторной коры. Под действием спонтанных возбуждающих команд другие нервные клетки переходят к активности, не дожидаясь осознанных сигналов; например, нейроны, управляющие движениями грудной клетки и диафрагмы при дыхании, подчиняются клеткам более высокого уровня, которые реагируют только на концентрацию О 2 и СО 2 в крови.
Судя по тому, что сегодня известно науке, межнейронные взаимодействия, происходящие в мозгу, можно в основном объяснить, исходя из возбуждающих и тормозных синаптических воздействий. Однако существуют и более сложные модифицирующие воздействия, которые имеют большое значение, так как они усиливают или уменьшают интенсивность ответной реакции нейрона на входные сигналы от различных других нейронов.
Рассмотрим модифицирующие медиаторные сигналы, представив себе, что они носят условный характер. Под термином «условный» подразумевается, что клетки реагируют на них только при определенных условиях, т.е. тогда, когда эти сигналы действуют в сочетании с другими возбуждающими или тормозными сигналами, приходящими по другим путям. Музыканты, например, могли бы считать условным действие педалей у фортепьяно - в том смысле, что для достижения какого-нибудь эффекта их нажатие должно сочетаться с другим действием. Просто нажимать на педали, не ударяя при этом по клавишам, бессмысленно. Звучание ноты изменяется лишь тогда, когда мы нажимаем одновременно и на педаль, и на клавишу. Многие нейронные сети, выполняющие условные функции, - это те, медиаторы которых играют важную роль в лечении депрессии, шизофрении и некоторых других психических расстройств (более подробно эти проблемы обсуждаются в гл. 9).
В заключение несколько слов о процессах, лежащих в основе различных изменений, производимых медиаторами в клетках, на которые они воздействуют. Эти изменения обусловлены ионными механизмами, связанными с электрической и химической регуляцией свойств мембраны. Возбудимость нейрона изменяется потому, что медиатор изменяет поток ионов, переходящих внутрь клетки или же из клетки наружу. Для того чтобы ионы могли проходить через мембрану, в ней должны быть отверстия. Это не просто дыры, а специальные крупные трубчатые белки, называемые «каналами». Некоторые из этих каналов специфичны для определенного иона - натрия, калия или кальция, например; другие не столь избирательны. Некоторые каналы могут открываться с помощью электрических команд (таких, как деполяризация мембраны при потенциале действия); другие открываются и закрываются под действием химических посредников.

Рис. 32. Схема адаптивных регуляторных процессов, используемых для поддержания нормальной синаптической передачи несмотря на изменения, вызываемые различными препаратами, а также, возможно, заболеваниями. Регулируется количество высвобождаемого или воспринимаемого медиатора. Слева - нормальное состояние. В середине - из-за недостаточного синтеза или сохранения медиатора постсинаптическая клетка увеличивает число рецепторов. Справа - при повышенном выбросе медиатора постсинаптическая клетка уменьшает число или эффективность рецепторов.
Полагают, что каждый химический посредник оказывает на клетки воздействие с помощью химически обусловленных изменений в ионной проницаемости. Определенные ионы и молекулы, используемые тем или иным медиатором, становятся поэтому химическим эквивалентом передаваемого сигнала.
Изменчивость нейронных функций
Как мы видели, нейрон должен успешно справляться с определенными задачами, чтобы функционировать как часть специфической нейронной сети. Медиатор, который он использует, должен передавать определенную информацию. У нейрона должны быть поверхностные рецепторы, с помощью которых он мог бы связывать медиатор в своих входных синапсах. Он должен иметь необходимые запасы энергии для «откачивания» избыточных ионов обратно через мембрану. Нейроны с длинными ветвящимися аксонами должны также транспортировать ферменты, медиаторы и другие молекулы из центральных участков цитоплазмы, где происходит их синтез, в отдаленные части дендритов и аксонов, где эти молекулы будут нужны. Как правило, скорость, с которой нейрон выполняет эти функции, зависит от массы его дендритной и аксонной систем и от общего уровня активности клетки.
Общая выработка энергии - метаболическая активность клетки - может изменяться в соответствии с требованиями межнейронных взаимодействий (рис. 32). Нейрон может повышать свою способность к синтезу и транспортировке специфических молекул в периоды усиленной деятельности. Точно так же при малой функциональной нагрузке нейрон может снизить уровень активности. Эта способность к регуляции фундаментальных внутриклеточных процессов позволяет нейрону гибко приспосабливаться к самым различным уровням активности.
Генетическая детерминация основных типов нейронных сетей
Для того чтобы мозг нормально функционировал, потоки нервных сигналов должны находить надлежащие маршруты среди клеток различных функциональных систем и межрегиональных объединений. В главе 1 мы получили некоторые элементарные сведения о сложном процессе построения и развития мозга. Однако до сих пор остается загадкой, каким образом аксоны и дендрита той или иной нервной клетки растут именно в том направлении, чтобы создавались специфические связи, необходимые для ее функционирования. Между тем тот факт, что конкретные молекулярные механизмы, лежащие в основе многих процессов онтогенеза, еще не раскрыты, не должен заслонять от нас другого, еще более поразительного факта-того, что из поколения в поколение в мозгу развивающихся животных действительно устанавливаются нужные связи. Исследования в области сравнительной нейроанатомии говорят о том, что по фундаментальному плану строения мозг очень мало изменился в процессе эволюции. Нейроны специализированного зрительного рецепторного органа - сетчатки - всегда соединяются с вторичными нейронами зрительной, а не слуховой или осязательной системы. В то же время первичные слуховые нейроны из специализированного органа слуха - улитки - всегда идут к вторичным нейронам слуховой системы, а не зрительной или обонятельной. Точно такая же специфичность связей характерна для любой системы мозга.
Высокая специфичность структуры мозга имеет важное значение. Общий диапазон связей для большинства нервных клеток, по-видимому, предопределен заранее , причем эта предопределенность касается тех клеточных свойств, которые ученые считают генетически контролируемыми . Набор генов, предназначенных для проявления в развивающейся нервной клетке, каким-то еще до конца не установленным образом определяет как будущий тип каждой нервной клетки, так и принадлежность ее к той или иной сети. Концепция генетической детерминированности приложима и ко всем остальным особенностям данного нейрона -например к используемому им медиатору, к размерам и форме клетки. Как внутриклеточные процессы, так и межнейронные взаимодействия определяются генетической специализацией клетки.
Три генетически детерминированных типа нервных сетей
Чтобы сделать концепцию генетической детерминации нейронных сетей более понятной, давайте уменьшим их число и представим себе, что наша нервная система состоит всего лишь из 9 клеток (см. рис. 33). Это абсурдное упрощение поможет нам увидеть сети трех основных типов, которые встречаются повсюду, - иерархические, локальные и дивергентные с одним входом . Хотя число элементов в сетях может быть различным, выделенные три типа могут служить основой для построения надежной классификационной схемы.
Иерархические сети . Наиболее распространенный тип межнейронных связей можно увидеть в главных сенсорных и двигательных путях. В сенсорных системах иерархическая организация носит восходящий характер; в нее включаются различные клеточные уровни, по которым информация поступает в высшие центры - от первичных рецепторов к вторичным вставочным нейронам, затем к третичным и т.д. Двигательные системы организованы по принципу нисходящей иерархии, где команды «спускаются» от нервной системы к мышцам: клетки, расположенные, фигурально говоря, «наверху», передают информацию специфическим моторным клеткам спинного мозга, а те в свою очередь - определенным группам мышечных клеток.
Иерархические системы обеспечивают очень точную передачу информации. В результате конвергенции (от лат converge - сходиться к одному центру) - когда несколько нейронов одного уровня контактируют с меньшим числом нейронов следующего уровня, или дивергенции (от лат. divergo - отклоняюсь, отхожу) - когда контакты устанавливаются с большим числом клеток следующего уровня, информация фильтруется и происходит усиление сигналов. Но, подобно любой цепи, иерархическая система не может быть сильнее своего самого слабого звена. Любая инактивация (от лат. in- - приставка, означающая отрицание) любого уровня, вызванная ранением, заболеванием, инсультом или опухолью, может вывести из строя всю систему. Конвергенция и дивергенция, однако, оставляют цепям некоторый шанс уцелеть даже при их серьезном повреждении, Если нейроны одного уровня будут частично уничтожены, сохранившиеся клетки смогут все-таки поддерживать функционирование сети.

Рис. 33. Нервная сеть из 9 клеток (схема). По периметру - нейроны соединены друг с другом в иерархическую цепь, типичную для сетей сенсорной и двигательной систем. В центре - дивергентная сеть с одним входом (клетки 5, 7, 8, 9), типичная для моноаминэргических систем, в которых один нейрон соединяется с большим числом мишеней. Слева - нейрон локальной сети (6), устанавливающий связи главным образом с клетками из своего ближайшего окружения.
Иерархические системы существуют, конечно, не только в сенсорных или двигательных путях. Тот же тип связей характерен для всех сетей, выполняющих какую-то специфическую функцию, т.е. для систем, которые мы назвали «альянсами» (гл. 1) и более подробно рассмотрим в последующих главах.
Локальные сети. Мы уже говорили о нейронах с короткими аксонами. Если у клетки короткий аксон, настолько короткий, что волнам электрической активности, можно сказать, некуда распространяться, очевидно, что задачи и сфера влияния такого нейрона должны быть весьма ограниченными. Нейроны локальных сетей действуют как фильтры, удерживая поток информации в пределах какого-то одного иерархического уровня. Они, по всей видимости, широко распространены во всех мозговых сетях.
Локальные сети могут оказывать на нейроны-мишени возбуждающее или тормозящее действие. Сочетание этих особенностей с дивергентным или ковергентным типом передачи на данном иерархическом уровне может еще более расширять, сужать или снова фокусировать поток информации.
Дивергентные сети с одним входом. В некоторых нервных сетях имеются скопления или слои нейронов, в которых один нейрон образует выходные связи с очень большим числом других клеток (в таких сетях дивергенция доведена до крайних пределов). Изучение сетей такого типа начато лишь недавно, и единственные места, где они встречаются (насколько нам сейчас известно), - это некоторые части среднего мозга и ствола мозга. Преимущества подобной системы в том, что она может оказывать влияние на множество нейронов сразу и иногда осуществлять связь со всеми иерархическими уровнями, нередко выходя за пределы специфических сенсорных, двигательных и других функциональных альянсов.
Поскольку сфера воздействия таких сетей не ограничена какой-либо системой с определенными функциями, дивергирующие пути этих сетей иногда называют неспецифическими . Однако ввиду того, что такие сети могут влиять на самые различные уровни и функции, они играют большую роль в интеграции многих видов деятельности нервной системы (см. гл. 4). Иными словами, такие системы выступают в роли организаторов и режиссеров массовых мероприятии, руководящих согласованными действиями больших групп людей. Кроме того, медиаторы, используемые в дивергентных системах с одним входом, - это медиаторы с «условным» действием: их эффект зависит от условии, в которых он осуществляется. Подобные воздействия весьма важны и для интегративных механизмов (лат. integratio - восстановление, восполнение, от integer - целый). Однако дивергентные сети такого типа составляют лишь небольшую часть всех нервных сетей.
Изменчивость генетически детерминированных типов сетей
Хотя общая картина связей специфических функциональных сетей удивительно сходна у всех, представителей одного вида, опыт каждой отдельной особи может оказывать дальнейшее влияние на межнейронные связи, вызывая в них индивидуальные изменения и корректируя их функцию.
Представим себе, например, что в мозгу большинства крыс каждый нейрон 3-го уровня в зрительной системе соединен примерно с 50 клетками-мишенями 4-го уровня - сравнительно небольшая дивергенция в системе, характеризующейся в остальном четкой иерархией. Теперь посмотрим, что произойдет, если крыса вырастет в полной темноте? Дефицит входной информации приведет к перестройке зрительной иерархии, так что каждый нейрон 3-го уровня будет контактировать только с 5 или 10 нейронами 4-го уровня вместо обычных 50. Однако, если мы рассмотрим нейроны 4-го уровня в микроскоп, мы убедимся, что у них нет недостатка во входных синапсах. Хотя зрительные нейроны 3-го уровня из-за малого числа связей передают информацию на 4-й уровень в ограниченном объеме, ее дефицит восполняется за счет других работающих сенсорных систем. У нашей крысы в доступном синаптическом пространстве 4-го уровня происходит процесс расширенной переработки слуховой и обонятельной информации.
Рассмотрим другой случай, где тот же эффект проявляется не столь резко. По некоторым данным, интенсивность межнейронной передачи сигналов может влиять на степень развития синаптических контактов между уровнями. Ряд ученых придерживается мнения, что некоторые формы памяти обусловлены изменениями в эффективности таких контактов. Эти изменения могут быть связаны как с микроструктурой (увеличение или уменьшение числа синапсов между клеткой А и клеткой Б), так и с действием медиаторов, участвующих в передаче сигналов (изменение количеств медиатора, синтезируемых и высвобождаемых одной клеткой, или степени реактивности другой клетки) (см. выше рис. 32). Эта тонкая регулировка локальных синаптических функций очень важна при некоторых заболеваниях мозга, о природе которых нам мало что известно (см. гл. 9). Малейшие изменения, происходящие на уровне синаптической активности, могли бы действительно вызвать аномалии поведения, но изменения эти настолько малы, что трудно установить, какова их роль на самом деле.
Нервные клетки не уникальны в своей способности к функциональным изменениям. Во многих других тканях клетки тоже могут изменяться, приспосабливаясь к нагрузке. Если мы возьмем небольшую пробу ткани из четырехглавой мышцы бедра у начинающего тяжелоатлета, а затем у него же после нескольких месяцев усиленной тренировки, то увидим, что каждое мышечное волокно содержит теперь сократимые фибриллы несколько большего размера и число этих фибрилл увеличилось. Слущивающиеся старые клетки вашей кожи и те, что выстилают желудочно-кишечный тракт, ежедневно заменяются новыми; эти клетки, однако, обладают способностью, которой нет у нейронов - они могут делиться. Нейроны генетически запрограммированы на синтез специфических молекул, с помощью которых работают синапсы, а также на образование весьма специфических связей, но не способны к делению. Представьте, что было бы, если бы нервные клетки стали делиться после образования синаптических связей. Как смогла бы при этом клетка распределить свои входные и выходные сигналы, чтобы сохранить прежние связи?
Хотя нейроны и не могут делиться, они обладают большей по сравнению с другими клетками способностью к адаптивной перестройке. Как показали эксперименты, в которых удаляли небольшой участок мозга, а затем в течение нескольких недель наблюдали за реакцией оставшихся частей, некоторые нервные клетки действительно могут регулировать степень своей связи с мишенями. Как правило, при повреждении некоторых синапсов одного нейрона другие, неповрежденные нейроны могут восполнить утраченные звенья цепи путем некоторого ускорения нормального процесса замены синапсов. Если две нервные клетки должны «общаться» более интенсивно, число связей между ними может возрастать за счет добавления новых синапсов при одновременном сохранении старых.
По-видимому, статичность макроскопического строения нервной системы заслонила от нас факт постоянного роста и отмирания связей. Существует даже мнение, что нейроны в нормальном состоянии все время образуют новые связи со своими мишенями. Как только новые синапсы сформировались, старые разрушаются. Такое замещение, вероятно, может компенсировать изнашивание связей в результате их длительной и непрерывной работы.
Хотя испытанное временем представление о том, что наш мозг не может регенерировать утраченные клетки, остается по-прежнему справедливым, исследования последних лет наводят на мысль, что здоровые нейроны обладают значительной структурной пластичностью. Этот более динамичный взгляд на изменчивость мозга открывает широкое поле для исследований; но прежде чем мы начнем понимать, как могут меняться синаптические связи, нам предстоит еще многое узнать.
Из книги Основы психофизиологии автора Александров Юрий2.7. Нейроны сетчатки Фоторецепторы сетчатки синаптически связаны с биполярными нервными клетками (см. рис. 4.2). При действии света уменьшается выделение медиатора из фоторецептора, что гиперполяризует мембрану биполярной клетки. От неё нервный сигнал передаётся на
Из книги Мозг рассказывает [Что делает нас людьми] автора Рамачандран Вилейанур С.3.4. Модулирующие нейроны В нервной системе выделена особая группа клеток – модулирующих нейронов, которые сами не вызывают реакции, но регулируют активность других нейронов. Они образуют контакты с другими нейронами типа «синапс на синапсе». Модулирующие нейроны
Из книги Эволюция человека. Книга 2. Обезьяны, нейроны и душа автора Марков Александр ВладимировичГлава 4 НЕЙРОНЫ, КОТОРЫЕ ОПРЕДЕЛИЛИ ЦИВИЛИЗАЦИЮ Даже когда мы одни, как часто с болью и удовольствием думаем мы о том, что другие думают о нас, об их воображаемом одобрении или порицании; все это следует из способности к сопереживанию, основного элемента социальных
Из книги Коннектом. Как мозг делает нас тем, что мы есть автора Сеунг СебастьянНейроны соревнуются за право запоминать Часто бывает так, что одни и те же важные сигналы, подлежащие запоминанию, принимаются одновременно очень многими нейронами. Нужно ли им всем участвовать в запоминании? На первый взгляд кажется, что это не слишком рационально. Ведь
Из книги автораГлава 4 Кругом одни нейроны Нервные импульсы да выработка нейротрансмиттеров – вот и всё. Что же, наше сознание выражается лишь этими физическими процессами, которые идут у нас в черепной коробке? Нейробиологи не сомневаются, что так и есть. Но большинство людей,
Из книги автораГлава 4. Кругом одни нейроны …позволяет ему делать и научные наблюдения… Quiroga et al., 2005.Даже фото Джулии Робертс… Эксперимент Фрида поражает, потому что был проделан на людях. Результаты поражают меньше, если вы знакомы с работами его предшественников, которые
14 Декабря 2017Нейроны – особая группа клеток организма, распространяющих информацию по всему телу. Используя электрические и химические сигналы, они помогают мозгу координировать все жизненно необходимые функции.
Если упростить, задачи нервной системы – собрать сигналы, поступающие из окружающей среды или из организма, оценить ситуацию, принять решение, как на них отреагировать (например, изменить частоту сердечных сокращений), а также подумать о происходящем и запомнить это. Основной инструмент для выполнения этих задач – нейроны, сплетенные по всему организму сложной сетью.
По средним оценкам, количество нейронов в головном мозге составляет 86 миллиардов, каждый из них связан еще с 1000 нейронов. Это создает невероятную сеть взаимодействия. Нейрон – основная единица нервной системы.
Нейроны (нервные клетки) составляют около 10% мозга, остальное – глиальные клетки и астроциты, функция которых заключается в поддержании и питании нейронов.
Как выглядит нейрон?
В строении нейрона можно выделить три части:
· Тело нейрона (сома) – получает информацию. Содержит ядро клетки.
· Дендриты – короткие отростки, принимающие информацию от других нейронов.
· Аксон – длинный отросток, несет информацию от тела нейрона в другие клетки. Чаще всего аксон оканчивается синапсом (контактом) с дендритами других нейронов.
Дендриты и аксоны называют нервными волокнами.
Аксоны сильно варьируют по длине, от нескольких миллиметров до метра и более. Самыми длинными являются аксоны спинномозговых ганглиев.
Типы нейронов
Классификацию нейронов можно провести по нескольким параметрам, например, по строению или выполняемой функции.
Типы нейронов в зависимости от функции:
· Эфферентные (двигательные) нейроны – несут информацию от центральной нервной системы (головного и спинного мозга) к клеткам других частей тела.
· Афферентные (чувствительные) нейроны – собирают информацию от всего организма и несут ее в центральную нервную систему.
· Вставочные нейроны – передают информацию между нейронами, чаще в пределах центральной нервной системы.
Как нейроны передают информацию?
Нейрон, получая информацию от других клеток, накапливает ее до тех пор, пока она не превысит определенный порог. После этого нейрон посылает по аксону электрический импульс – потенциал действия.
Потенциал действия формируется движением электрически заряженных частиц через мембрану аксона.
В состоянии покоя электрический заряд внутри нейрона отрицательный относительно окружающей его межклеточной жидкости. Эта разница называется мембранным потенциалом. Обычно он составляет 70 милливольт.
Когда тело нейрона получает достаточно заряда, и он «выстреливает», в соседнем участке аксона происходит деполяризация – мембранный потенциал быстро растет, а затем падает примерно за 1/1000 секунды. Этот процесс запускает деполяризацию соседнего участка аксона, и так далее, пока импульс не пройдет по всей длине аксона. После процесса деполяризации наступает гиперполяризация – кратковременное состояние отдыха, в этот момент передача импульса невозможна.

Потенциал действия чаще всего генерируют ионы калия (К+) и натрия (Na+), которые по ионным каналам перемещаются из межклеточной жидкости внутрь клетки и обратно, меняя заряд нейрона и делая его сначала положительным, а затем снижая его.
Потенциал действия обеспечивает работу клетки по принципу «все или ничего», то есть импульс или передается, или нет. Слабые сигналы будут накапливаться в теле нейрона до тех пор, пока их заряда не будет достаточно для передачи по отросткам.
Миелин
Миелин – это белое густое вещество, покрывающее большинство аксонов. Это покрытие обеспечивает электроизоляцию волокна и повышает скорость прохождения импульса по нему.

Миелинизированное волокно в сравнении с немиелинизированным.
Миелин вырабатывается шванновскими клетками на периферии и олигодендроцитами в центральной нервной системе. По ходу волокна миелиновая оболочка прерывается – это перехваты Ранвье. Потенциал действия перемещается от перехвата к перехвату, что обеспечивает быструю передачу импульса.
Такое распространенное и серьезное заболевание, как рассеянный склероз, вызвано разрушением миелиновой оболочки.
Как работают синапсы
Нейроны и ткани, которым они передают импульс, физически не соприкасаются, между клетками всегда существует пространство – синапс.
В зависимости от способа передачи информации, синапсы могут быть химическими и электрическими.
Химический синапсПосле того как сигнал, передвигаясь по отростку нейрона, достигает синапса, происходит высвобождение химических веществ – нейромедиаторов (нейротрансмиттеров) в пространство между двумя нейронами. Это пространство называют синаптической щелью.

Схема строения химического синапса.
Нейромедиатор из передающего (пресинаптического) нейрона, попадая в синаптическую щель, взаимодействует с рецепторами на мембране принимающего (постсинаптического) нейрона, запуская целую цепь процессов.
Виды химических синапсов:
· глютаматэргический – медиатором является глютаминовая кислота, обладает возбуждающим эффектом на синапс;
· ГАМК-эргический – медиатором является гамма-аминомасляная кислота (ГАМК), обладает тормозящим эффектом на синапс;
· холинергический – медиатором является ацетилхолин, осуществляет нервно-мышечную передачу информации;
· адренергический – медиатором является адреналин.
Электрические синапсы
Электрические синапсы встречаются реже, распространены в центральной нервной системе. Клетки сообщаются посредством особых белковых каналов. Пресинаптическая и постсинаптическая мембраны в электрических синапсах расположены близко друг к другу, поэтому импульс способен проходить непосредственно от клетки к клетке.
Скорость передачи импульса по электрическим синапсам гораздо выше, чем по химическим, поэтому они расположены преимущественно в тех отделах, где необходима быстрая реакция, например, отвечающих за защитные рефлексы.
Еще одно отличие двух типов синапсов в направлении передачи информации: если химические синапсы могут передавать импульс только в одном направлении, то электрические в этом смысле универсальны.
Заключение
Нейроны – это, пожалуй, самые необычные клетки организма. Каждое действие, которое осуществляет тело человека, обеспечивается работой нейронов. Сложная нейронная сеть формирует личность и сознание. Они отвечают как за самые примитивные рефлексы, так и за самые сложные процессы, связанные с мышлением.
С моим видением того как работает мозг и каковы возможные пути создания искусственного интеллекта. За прошедшее с тех пор время удалось существенно продвинуться вперед. Что-то получилось глубже понять, что-то удалось смоделировать на компьютере. Что приятно, появились единомышленники, активно участвующие в работе над проектом.
В настоящем цикле статей планируется рассказать о той концепции интеллекта над которой мы сейчас работаем и продемонстрировать некоторые решения, являющиеся принципиально новыми в сфере моделирования работы мозга. Но чтобы повествование было понятным и последовательным оно будет содержать не только описание новых идей, но и рассказ о работе мозга вообще. Какие-то вещи, особенно в начале, возможно покажутся простыми и общеизвестными, но я бы советовал не пропускать их, так как они во многом определяют общую доказательность повествования.
Общее представление о мозге
Нервные клетки, они же нейроны, вместе со своими волокнами, передающими сигналы, образуют нервную систему. У позвоночных основная часть нейронов сосредоточена в полости черепа и позвоночном канале. Это называется центральной нервной системой. Соответственно, выделяют головной и спинной мозг как ее составляющие.
Спинной мозг собирает сигналы от большинства рецепторов тела и передает их в головной мозг. Через структуры таламуса они распределяются и проецируются на кору больших полушарий головного мозга.
Кроме больших полушарий обработкой информации занимается еще и мозжечок, который, по сути, является маленьким самостоятельным мозгом. Мозжечок обеспечивает точную моторику и координацию всех движений.
Зрение, слух и обоняние обеспечивают мозг потоком информации о внешнем мире. Каждая из составляющих этого потока, пройдя по своему тракту, также проецируется на кору. Кора – это слой серого вещества толщиной от 1.3 до 4.5 мм, составляющий наружную поверхность мозга. За счет извилин, образованных складками, кора упакована так, что занимает в три раза меньшую площадь, чем в расправленном виде. Общая площадь коры одного полушария – приблизительно 7000 кв.см.
В итоге все сигналы проецируются на кору. Проекция осуществляется пучками нервных волокон, которые распределяются по ограниченным областям коры. Участок, на который проецируется либо внешняя информация, либо информация с других участков мозга образует зону коры. В зависимости от того, какие сигналы на такую зону поступают, она имеет свою специализацию. Различают моторную зону коры, сенсорную зону, зоны Брока, Вернике, зрительные зоны, затылочную долю, всего около сотни различных зон.

В вертикальном направлении кору принято делить на шесть слоев. Эти слои не имеют четких границ и определяются по преобладанию того или иного типа клеток. В различных зонах коры эти слои могут быть выражены по-разному, сильнее или слабее. Но, в общем и целом, можно говорить о том, что кора достаточно универсальна, и предполагать, что функционирование разных ее зон подчиняется одним и тем же принципам.

Слои коры
По афферентным волокнам сигналы поступают в кору. Они попадают на III, IV уровень коры, где распределяются по близлежащим к тому месту, куда попало афферентное волокно, нейронам. Большая часть нейронов имеет аксонные связи в пределах своего участка коры. Но некоторые нейроны имеют аксоны, выходящие за ее пределы. По этим эфферентным волокнам сигналы идут либо за пределы мозга, например, к исполнительным органам, или проецируются на другие участки коры своего или другого полушария. В зависимости от направления передачи сигналов эфферентные волокна принято делить на:
- ассоциативные волокна, которые связывают отдельные участки коры одного полушария;
- комиссуральные волокна, которые соединяют кору двух полушарий;
- проекционные волокна, которые соединяют кору с ядрами низших отделов центральной нервной системы.
Можно представить себе кору головного мозга как большое полотно, раскроенное на отдельные зоны. Картина активности нейронов каждой из зон кодирует определенную информацию. Пучки нервных волокон, образованные аксонами, выходящими за пределы своей зоны коры, формируют систему проекционных связей. На каждую из зон проецируется определенная информация. Причем на одну зону может поступать одновременно несколько информационных потоков, которые могут приходить как с зон своего, так и противоположного полушария. Каждый поток информации похож на своеобразную картинку, нарисованную активностью аксонов нервного пучка. Функционирование отдельной зоны коры – это получение множества проекций, запоминание информации, ее переработка, формирование собственной картины активности и дальнейшая проекция информации, получившейся в результате работы этой зоны.
Существенный объем мозга – это белое вещество. Оно образовано аксонами нейронов, создающими те самые проекционные пути. На рисунке ниже белое вещество можно увидеть как светлое заполнение между корой и внутренними структурам мозга.

Распределение белого вещества на фронтальном срезе мозга
Используя диффузную спектральную МРТ, удалось отследить направление отдельных волокон и построить трехмерную модель связанности зон коры (проект Connectomics (Коннектом)).
Представление о структуре связей хорошо дают рисунки ниже (Van J. Wedeen, Douglas L. Rosene, Ruopeng Wang, Guangping Dai, Farzad Mortazavi, Patric Hagmann, Jon H. Kaas, Wen-Yih I. Tseng, 2012).

Вид со стороны левого полушария

Вид сзади

Вид справа
Кстати, на виде сзади отчетливо видна асимметрия проекционных путей левого и правого полушария. Эта асимметрия во многом и определяет различия в тех функциях, которые приобретают полушария по мере их обучения.
Нейрон
Основа мозга – нейрон. Естественно, что моделирование мозга с помощью нейронных сетей начинается с ответа на вопрос, каков принцип его работы.
В основе работы реального нейрона лежат химические процессы. В состоянии покоя между внутренней и внешней средой нейрона существует разность потенциалов – мембранный потенциал, составляющий около 75 милливольт. Он образуется за счет работы особых белковых молекул, работающих как натрий-калиевые насосы. Эти насосы за счет энергии нуклеотида АТФ гонят ионы калия внутрь, а ионы натрия - наружу клетки. Поскольку белок при этом действует как АТФ-аза, то есть фермент, гидролизующий АТФ, то он так и называется - «натрий-калиевая АТФ-аза». В результате нейрон превращается в заряженный конденсатор с отрицательным зарядом внутри и положительным снаружи.

Схема нейрона (Mariana Ruiz Villarreal)
Поверхность нейрона покрыта ветвящимися отростками – дендритами. К дендритам примыкают аксонные окончания других нейронов. Места их соединений называются синапсами. Посредством синаптического взаимодействия нейрон способен реагировать на поступающие сигналы и при определенных обстоятельствах генерировать собственный импульс, называемый спайком.
Передача сигнала в синапсах происходит за счет веществ, называемых нейромедиаторами. Когда нервный импульс по аксону поступает в синапс, он высвобождает из специальных пузырьков молекулы нейромедиатора, характерные для этого синапса. На мембране нейрона, получающего сигнал, есть белковые молекулы – рецепторы. Рецепторы взаимодействуют с нейромедиаторами.

Химический синапс
Рецепторы, расположенные в синаптической щели, являются ионотропными. Это название подчеркивает тот факт, что они же являются ионными каналами, способными перемещать ионы. Нейромедиаторы так воздействуют на рецепторы, что их ионные каналы открываются. Соответственно, мембрана либо деполяризуется, либо гиперполяризуется – в зависимости от того, какие каналы затронуты и, соответственно, какого типа этот синапс. В возбуждающих синапсах открываются каналы, пропускающие катионы внутрь клетки, - мембрана деполяризуется. В тормозных синапсах открываются каналы, проводящие анионы, что приводит к гиперполяризации мембраны.
В определенных обстоятельствах синапсы могут менять свою чувствительность, что называется синаптической пластичностью. Это приводит к тому, что синапсы одного нейрона приобретают различную между собой восприимчивость к внешним сигналам.
Одновременно на синапсы нейрона поступает множество сигналов. Тормозящие синапсы тянут потенциал мембраны в сторону накопления заряда внутри клети. Активирующие синапсы, наоборот, стараются разрядить нейрон (рисунок ниже).

Возбуждение (A) и торможение (B) ганглиозной клетки сетчатки (Николлс Дж., Мартин Р., Валлас Б., Фукс П., 2003)
Когда суммарная активность превышает порог инициации, возникает разряд, называемый потенциалом действия или спайком. Спайк – это резкая деполяризация мембраны нейрона, которая и порождает электрический импульс. Весь процесс генерации импульса длится порядка 1 миллисекунды. При этом ни продолжительность, ни амплитуда импульса не зависят от того, насколько были сильны вызвавшие его причины (рисунок ниже).

Регистрация потенциала действия ганглиозной клетки (Николлс Дж., Мартин Р., Валлас Б., Фукс П., 2003)
После спайка ионные насосы обеспечивают обратный захват нейромедиатора и расчистку синаптической щели. В течение рефрактерного периода, наступающего после спайка, нейрон не способен порождать новые импульсы. Продолжительность этого периода определяет максимальную частоту генерации, на которую способен нейрон.
Спайки, которые возникают как следствие активности на синапсах, называют вызванными. Частота следования вызванных спайков кодирует то, насколько хорошо поступающий сигнал соответствует настройке чувствительности синапсов нейрона. Когда поступающие сигналы приходятся именно на чувствительные синапсы, активирующие нейрон, и этому не мешают сигналы, приходящие на тормозные синапсы, то реакция нейрона максимальна. Образ, который описывается такими сигналами, называют характерным для нейрона стимулом.
Конечно, представление о работе нейронов не стоит излишне упрощать. Информация между некоторыми нейронами может передаваться не только спайками, но и за счет каналов, соединяющих их внутриклеточное содержимое и передающих электрический потенциал напрямую. Такое распространение называется градуальным, а само соединение называется электрическим синапсом. Дендриты в зависимости от расстояния до тела нейрона делятся на проксимальные (близкие) и дистальные (удаленные). Дистальные дендриты могут образовывать секции, работающие как полуавтономные элементы. Помимо синаптических путей возбуждения есть внесинаптические механизмы, вызывающие метаботропные спайки. Кроме вызванной активности существует еще и спонтанная активность. И наконец, нейроны мозга окружены глиальными клетками, которые также оказывают существенное влияние на протекающие процессы.
Долгий путь эволюции создал множество механизмов, которые используются мозгом в своей работе. Некоторые из них могут быть поняты сами по себе, смысл других становится ясен только при рассмотрении достаточно сложных взаимодействий. Поэтому не стоит воспринимать сделанное выше описание нейрона как исчерпывающее. Чтобы перейти к более глубоким моделям, нам необходимо сначала разобраться с «базовыми» свойствами нейронов.
В 1952 году Аланом Ллойдом Ходжкином и Эндрю Хаксли были сделаны описания электрических механизмов, которые определяют генерацию и передачу нервного сигнала в гигантском аксоне кальмара (Hodgkin, 1952). Что было оценено Нобелевской премией в области физиологии и медицины в 1963 году. Модель Ходжкина – Хаксли описывает поведение нейрона системой обыкновенных дифференциальных уравнений. Эти уравнения соответствуют автоволновому процессу в активной среде. Они учитывают множество компонент, каждая из которых имеет свой биофизический аналог в реальной клетке (рисунок ниже). Ионные насосы соответствуют источнику тока I p . Внутренний липидный слой клеточной мембраны образует конденсатор с емкостью C m . Ионные каналы синаптических рецепторов обеспечивают электрическую проводимость g n , которая зависит от подаваемых сигналов, меняющихся со временем t, и общей величины мембранного потенциала V. Ток утечки мембранных пор создает проводник g L . Движение ионов по ионным каналам происходит под действием электрохимических градиентов, которым соответствуют источники напряжения с электродвижущей силой E n и E L .

Основные компоненты модели Ходжкина - Хаксли
Естественно, что при создании нейронных сетей возникает желание упростить модель нейрона, оставив в ней только самые существенные свойства. Наиболее известная и популярная упрощенная модель – это искусственный нейрон Маккалока - Питтса, разработанный в начале 1940-х годов (Маккалох Дж., Питтс У., 1956).
Формальный нейрон Маккалока - Питтса
На входы такого нейрона подаются сигналы. Эти сигналы взвешенно суммируются. Далее к этой линейной комбинации применяется некая нелинейная функция активации, например, сигмоидальная. Часто как сигмоидальную используют логистическую функцию:![]()

Логистическая функция
В этом случае активность формального нейрона записывается как

В итоге такой нейрон превращается в пороговый сумматор. При достаточно крутой пороговой функции сигнал выхода нейрона – либо 0, либо 1. Взвешенная сумма входного сигнала и весов нейрона – это свертка двух образов: образа входного сигнала и образа, описываемого весами нейрона. Результат свертки тем выше, чем точнее соответствие этих образов. То есть нейрон, по сути, определяет, насколько подаваемый сигнал похож на образ, записанный на его синапсах. Когда значение свертки превышает определенный уровень и пороговая функция переключается в единицу, это можно интерпретировать как решительное заявление нейрона о том, что он узнал предъявляемый образ.
Реальные нейроны действительно неким образом похожи на нейроны Маккалока - Питтса. Амплитуды их спайков не зависит от того, какие сигналы на синапсах их вызвали. Спайк, либо есть, либо его нет. Но реальные нейроны реагируют на стимул не единичным импульсом, а импульсной последовательностью. При этом частота импульсов тем выше, чем точнее узнан характерный для нейрона образ. Это означает, что если мы построим нейронную сеть из таких пороговых сумматоров, то она при статичном входном сигнале хотя и даст какой-то выходной результат, но этот результат будет далек от воспроизведения того, как работают реальные нейроны. Для того чтобы приблизить нейронную сеть к биологическому прототипу, нам понадобится моделировать работу в динамике, учитывая временные параметры и воспроизводя частотные свойства сигналов.
Но можно пойти и другим путем. Например, можно выделить обобщенную характеристику активности нейрона, которая соответствует частоте его импульсов, то есть количеству спайков за определенный промежуток времени. Если перейти к такому описанию, то можно представить нейрон как простой линейный сумматор.

Линейный сумматор
Сигналы выхода и, соответственно, входа для таких нейронов уже не являются дихатомичными (0 или 1), а выражаются некой скалярной величиной. Функция активации тогда записывается как
Линейный сумматор не стоит воспринимать как что-то принципиально иное по сравнению с импульсным нейроном, просто он позволяет при моделировании или описании перейти к более длинным временным интервалам. И хотя импульсное описание более корректно, переход к линейному сумматору во многих случаях оправдан сильным упрощением модели. Более того, некоторые важные свойства, которые трудно разглядеть в импульсном нейроне, вполне очевидны для линейного сумматора.